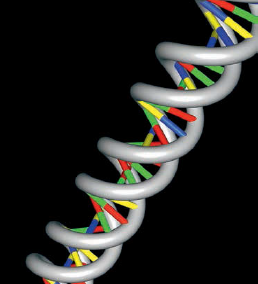
A estrutura básica do DNA
As ligações de hidrogênio estão presentes em cada um dos pareamentos dos nucleotídeos, mas podem ser rompidas, o que determina o despareamento das duas hélices da molécula. O calor é um dos fatores que podem contribuir para esse rompimento, chamado desnaturação. Os trechos do DNA com maior concentração de pares citosina-guanina são mais resistentes à desnaturação, pois têm três ligações de hidrogênio.
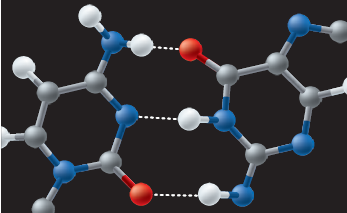
Os trechos ricos em pareamentos adenina-timina desnaturam-se mais facilmente, pois estabelecem apenas duas ligações de hidrogênio em cada pareamento.
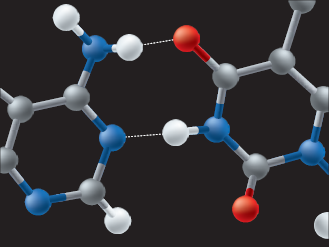
de hidrogênio (em branco) de um lado e os de nitrogênio (em azul) ou de oxigênio (em vermelho) de outro determina o aparecimento de ligações de hidrogênio entre eles (representada pelo pontilhado branco), que estabiliza a posição relativa das bases (A=T).
Em geral, a desnaturação do DNA pode ser revertida facilmente, restabelecendo os pareamentos da molécula.
Observe a figura abaixo.

Nela é possível identificar a molécula de pentose (desoxirribose), as moléculas de fosfato e as bases hidrogenadas. Note, ainda, a posição dos átomos de oxigênio (cor vermelha) no anel da molécula de desoxirribose. A informação genética está armazenada nessa sequência de bases nitrogenadas.
presas as bases nitrogenadas devidamente pareadas: guaninas (verde) com citosinas (vermelho) e timinas (azul) com adeninas (amarelo).
Assim como a sequência de letras dá significado às palavras em nossa escrita, a sequência de bases nitrogenadas constitui uma informação que a célula é capaz de decifrar.
A informação genética inscrita no DNA revela a sequência dos aminoácidos que farão parte da molécula de proteínas, ou seja, a estrutura primária da proteína. Você viu que essa sequência de aminoácidos é muito precisa e rigorosa, e que, às vezes, a troca de um único aminoácido muda as propriedades funcionais da proteína, como no caso da hemoglobina produzida na anemia falciforme.

Você viu que vinte aminoácidos fazem parte das proteínas dos organismos. Como o DNA não possui vinte bases nitrogenadas, já se supunha que a informação genética residia na ordem de disposição das bases nitrogenadas ao longo da molécula de DNA. Outra característica importante do modelo de Watson e Crick foi imaginar que o DNA formasse uma cadeia complementar, compondo uma dupla hélice. Watson e Crick perceberam que essa estrutura dupla permitiria armazenar a informação com segurança. Além disso, seria possível explicar como o material genético é duplicado.
Cada uma das duas fitas poderia servir de molde para a formação de duas novas fitas, que seriam duplicadas mantendo a integridade da informação armazenada na sequência de bases nitrogenadas.

Watson e Crick, ao propor a estrutura em dupla hélice, registraram que tinham percebido como a estrutura em forma de dupla fita permitia duplicar a molécula, formando duas moléculas iguais, exatamente com a mesma sequência de bases nitrogenadas, cada qual composta de uma hélice antiga e de outra recém-formada.
Assim, eles levantaram a hipótese de que a replicação do DNA deveria ser semiconservativa. O modelo semiconservativo de duplicação do DNA foi logo colocado em teste por meio da marcação do DNA com elementos radioativos. Em 1957 foram publicados resultados nos quais os cromossomos das células da raiz de uma planta foram marcados com uma base nitrogenada contendo uma forma de hidrogênio radioativo. Colocando essas células em uma cultura com bases nitrogenadas sem radioatividade, percebeu-se que todos os cromossomos das células-filhas da primeira geração permaneciam com radioatividade. Na segunda geração, apenas metade dos cromossomos das células-filhas mantinham radioatividade. Esse era justamente o resultado esperado pelo modelo semiconservativo.
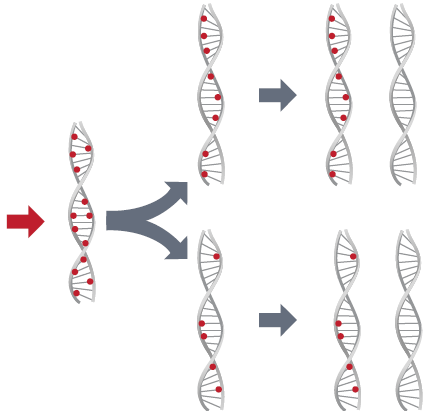
No ano seguinte, um experimento com bactérias chegou aos mesmos resultados, mas com uma técnica mais sofisticada. Os norte-americanos Matthew Meselson e Franklin Stahl cultivaram bactérias em meio com uma única fonte de nitrogênio marcado, que tinha a vantagem de manter a radioatividade por muito mais tempo do que o hidrogênio. As bactérias com radioatividade foram então transferidas para um meio com nitrogênio comum. A cada geração o DNA era purificado e era feita uma medida muito precisa da quantidade de cada forma de nitrogênio. As bactérias da primeira geração, mesmo crescendo em meio livre de radioatividade, mantinham a metade da radioatividade do lote original. Apenas na geração seguinte apareciam bactérias livres de radioatividade. O experimento ficou conhecido como Meselson-Stahl, confirmava resultados anteriores e convencia a comunidade científica de que o modelo semiconservativo descrevia corretamente a duplicação do DNA.
Referências:
Bizzo, Nélio Novas bases da biologia / Nélio Bizzo. – 2. ed. – São Paulo : Ática, 2013. Conteúdo : v. 1 Células, organismos e populações – v. 2 Biodiversidade – v.3 Corpo humano, genes e ambiente
Was this helpful?
0 / 0